




| | | | |
 |  |  |  |  |  |  |

The diagram above is the distribution in time excluding the European E1b-V13. This gives a better view on the different population jumps of the Afroasiatic branches.
Several aspects in the time estimates are uncertain. In the Indo-European data, the moment of migration of R1b to Anatolia is the clearest strong migration. The Y-DNA yfull time estimates indicate 5300 ybp, which is about 20 percent larger than the glottochronology estimate of Holman et al. The time estimate of N in the Ural gives about 4000 ybp for the Uralic languages. This is about 25 percent larger than the estimate of Holman et al. The first Niger-Congo expansion that we see is 8900 ybp. The next expansion of this branch (followed by more expansions) was near 6100 ybp. If we use the first expansion it would be about 40 percent larger. If we use the E-L1250 (in E-V22; 7900 ybp) population growth as the basic population jump for the Afroasiatic languages, it would result in a correction of 30 percent.
If we have a look at the relative population sizes of E-M35 (excluding the Indo-European E-V13) between 8000-2000 ybp, we see a few moments of population jumps in specific branches. We see the E-V22 branch expanding near 8000 ybp, and continuing the population growth at 7500 ybp in E-V22 and also in E-M84, probably originating in the Egypt and the start of the Afroasiatic languages. The Egyptian language is probably directly related to the population jump near 7500ybp. The E-M84 has a second expansion moment at 5500 ybp (see E-S9747), which was probably the start of the Semitic languages in the Levant. The last expansion is the E-M81 expansion in the Maghreb, which is probably the start of the Berber languages. If the identification of the last two are correct, they have a correction of 65 (3301ybp) and 25 percent (1733) respectively. The uncertainty of time estimates is probably in the glottochronology, which is know to be uncertain. The estimate of E-S9747 (5600 ybp) corresponds with the Kish civilization which has the oldest traces of a Semitic language.
The Cushitic languages have its distribution near Somalia. I looked at the Y-DNA characteristics in the Cushitic language peoples in yfull and ftdna. Their largest shared Y-DNA branch is E1b-V12-V32-Y18629. Their shared ancestor has three descending branches, and one them (E-Y18637) has only one SNP and four descending branches. This combination is a population jump and occured at 2700ybp. The combination of a population jump, a majority in a language region makes it likely that this was the start of the Cushic languages. It follows the pattern that we saw before. The Holman estimate is 4734ybp, which does not fit. This can only fit if the E1b-Y18629 branch represents a branch of a more recent language within the Cushitic languages.
The Chadic languages have a Y-DNA heritage of R-Y7771. This branch split has at present 4 descending branches (5200 ybp) and R-V69 has at present 5 descending branches (4500 ybp). The presence of a few Arab branches is consistent with the arrival of Arabs in the Sahara and Sahel regions and exchanging people in both directions. The explanation of Cruciani et al (2010) and Shriner et al (2018) are unlikely to tell the complete story. The Shriner scenario does not explain the many old branches of R1b-V88. It seems likely that R1b-V88 played a role in the distribution of the Chadic language, so starting 5200 ybp. Whether they received their language in the Sahel or outside Africa is unclear. An explanation of an arrival from the north (samples in Italy and Serbia) passing the Green Sahara or the Middle East are both possible. It seems likely that their received the language from the female lines in the Africa and expanded it as a R1b-V88 with this Chadic language after 5200ybp. The Holman estimate is 4826ybp, so a correction of 8 percent is required.
It seems likely that the 7500ybp population jump of E-V22 and E-M84 had the largest impact on the Afroasiatic languages. If we follow the diagram of Lipinski (2001) the easiest fit would give a shared ancestor of Chadic and Egyptian before the population jumps of 7500ybp in the Green Sahara. I saw no indication how the Cushitic language would fit into this scheme. One can think of an earlier branch split or a later but faster language change in a smaller group.
This population spread was probably the main contributor to the population that started the civilization of ancient Egypt.
Five Natufians (Levant, 10 BCE) were analyzed in the Laradis et al paper (2016). The DNA is old; two were reported with SNPs equal to E-Z830. A third was in a different E-branch. It shows that haplogroup E was present in this area before the population jumps of E-V22.
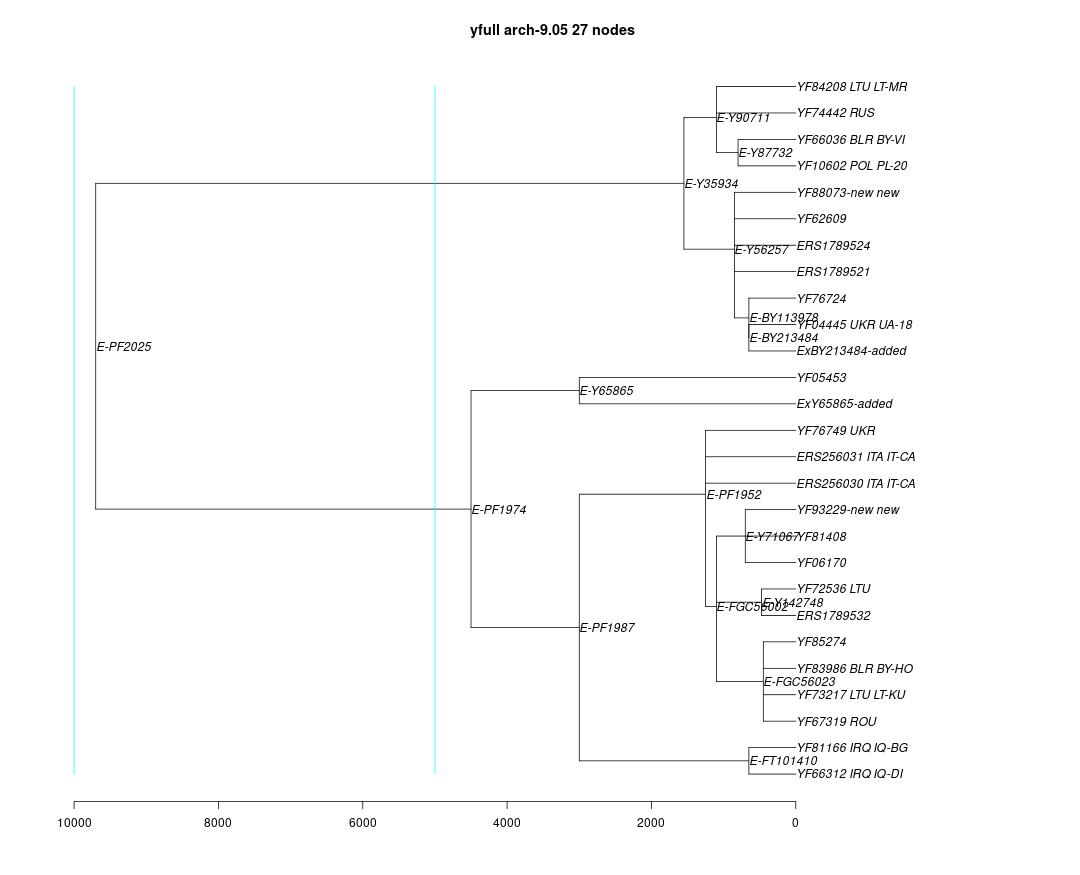
link to phylogenetic tree: E-PF2025
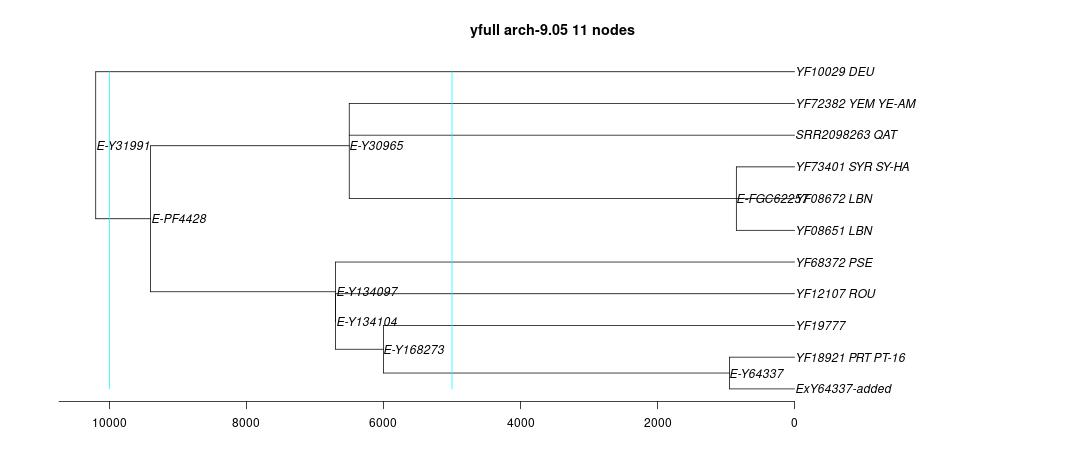
link to phylogenetic tree: E-Y31991
link to phylogenetic tree: E-M84
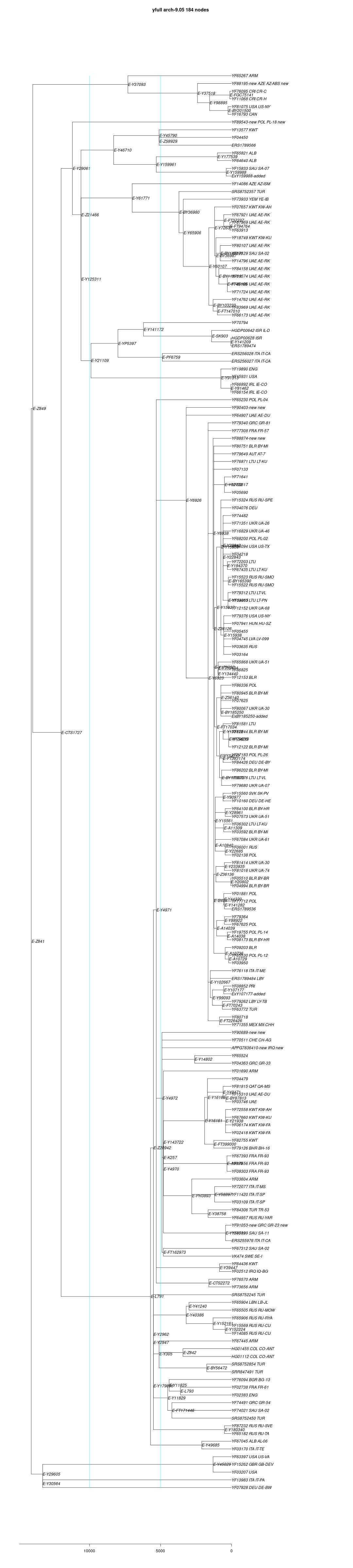
link to phylogenetic tree: E-Z841
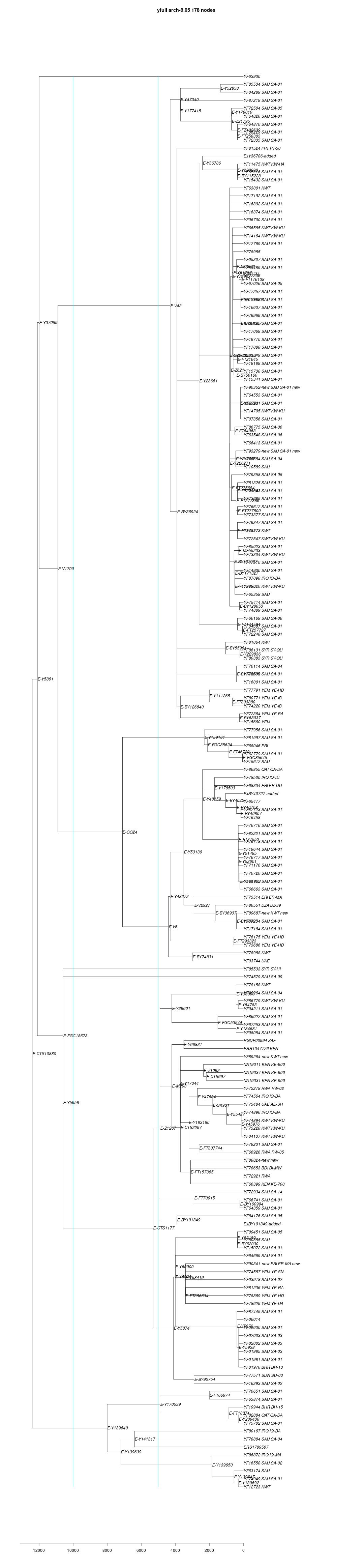
link to phylogenetic tree: E-CTS10880
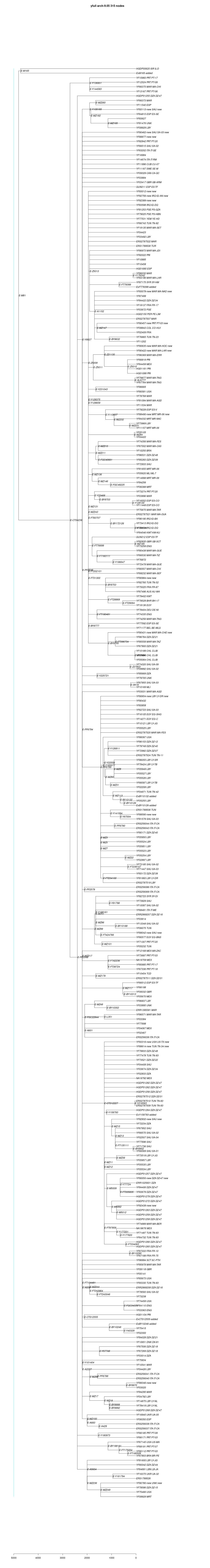
link to phylogenetic tree: E-M81
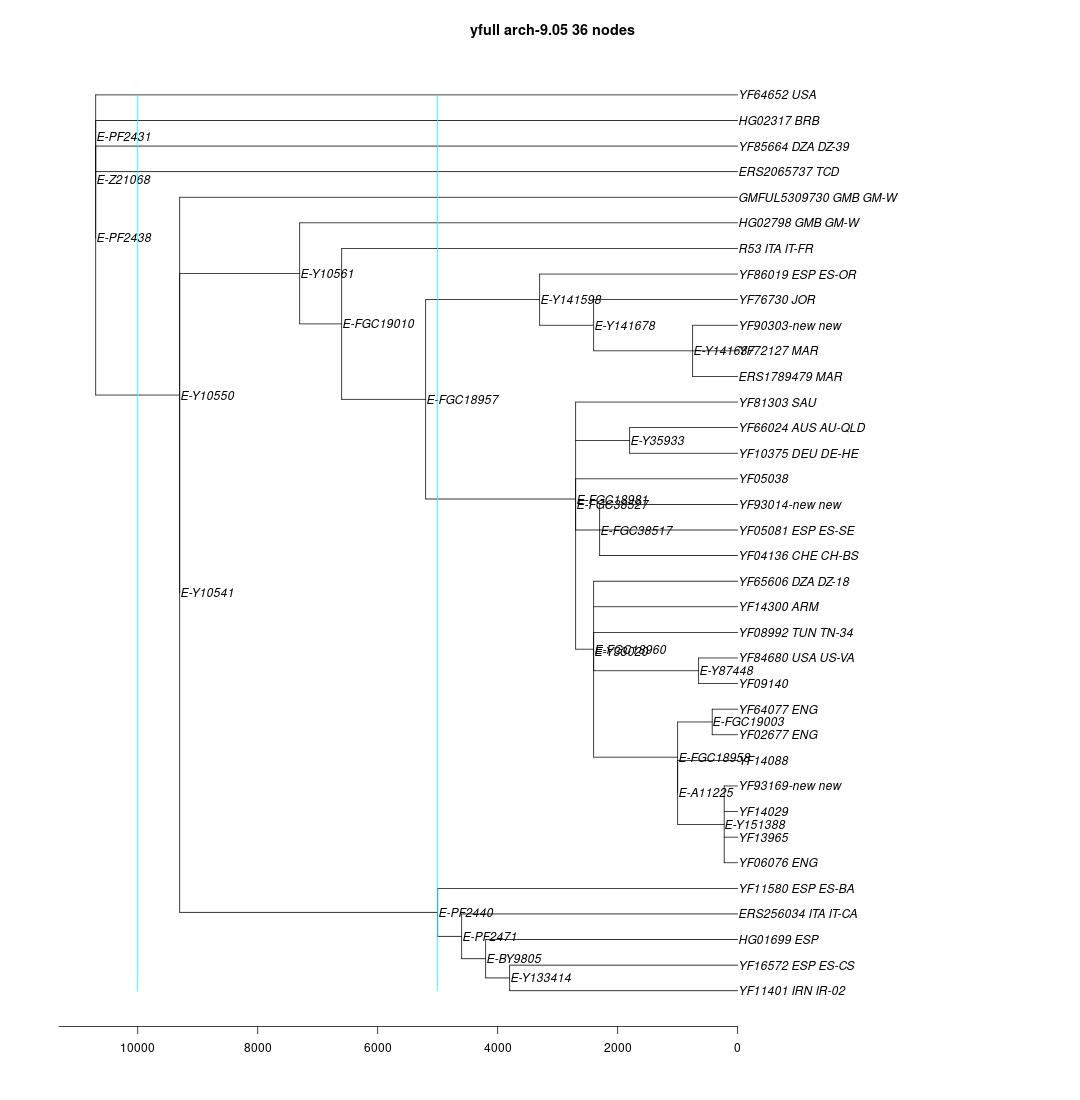
link to phylogenetic tree: E-PF2431
link to phylogenetic tree: E-L618
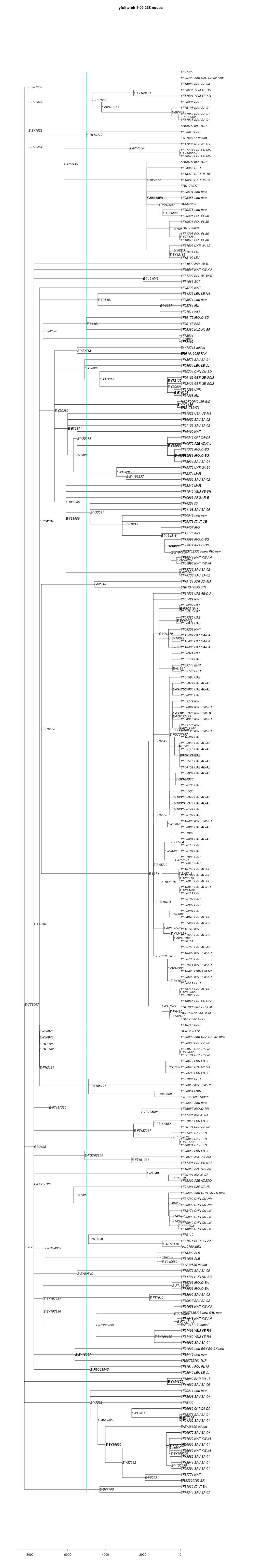
link to phylogenetic tree: E-V22
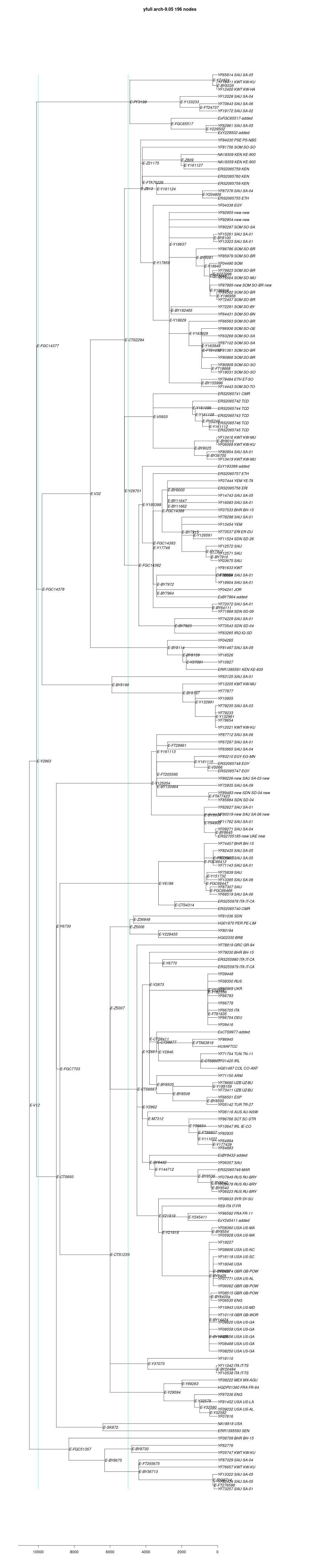
link to phylogenetic tree: E-V12
link to phylogenetic tree: E-Z22639
yfull-links: E - E-M5479 - E-P147 - E-P177 - E-M215 - E-M35 - E-Z827 - E-Z830 - E-PF1962 - E-PF2025
yfull-links: E - E-M5479 - E-P147 - E-P177 - E-M215 - E-M35 - E-Z827 - E-Z830 - E-PF1962 - E-M123 - E-Y31991
yfull-links: E - E-M5479 - E-P147 - E-P177 - E-M215 - E-M35 - E-Z827 - E-Z830 - E-PF1962 - E-M123 - E-M34 - E-M84
yfull-links: E - E-M5479 - E-P147 - E-P177 - E-M215 - E-M35 - E-Z827 - E-Z830 - E-PF1962 - E-M123 - E-M34 - E-Z841
yfull-links: E - E-M5479 - E-P147 - E-P177 - E-M215 - E-M35 - E-Z827 - E-Z830 - E-CTS10880
yfull-links: E - E-M5479 - E-P147 - E-P177 - E-M215 - E-M35 - E-Z827 - E-L19 - E-M81
yfull-links: E - E-M5479 - E-P147 - E-P177 - E-M215 - E-M35 - E-Z827 - E-L19 - E-PF2431
yfull-links: E - E-M5479 - E-P147 - E-P177 - E-M215 - E-M35 - E-L539 - E-M78 - E-Z1919 - E-L618
yfull-links: E - E-M5479 - E-P147 - E-P177 - E-M215 - E-M35 - E-L539 - E-M78 - E-Z1919 - E-V22
yfull-links: E - E-M5479 - E-P147 - E-P177 - E-M215 - E-M35 - E-L539 - E-M78 - E-Z1902 - E-V12
yfull-links: E - E-M5479 - E-P147 - E-P177 - E-M215 - E-M35 - E-L539 - E-M78 - E-Z1902 - E-Z22639


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}